- 大分類
-
- 血管
- 小分類
-
- 機構
血管新生 angiogenesis
解説
【概要】
【機序】
既存血管では、血管内皮細胞-壁細胞間の接着が強固であり、内皮細胞は基底膜側から血管外には移動に抑制がかかっている。内皮細胞の無血管野への移動からはじまる血管新生の開始のためには、この細胞間接着を抑制する必要がある。壁細胞から分泌されるアンジオポエチン1(Ang1)が内皮細胞上の受容体型チロシンキナーゼTie2を活性化して内皮-壁細胞間の細胞接着を誘導しているが、血管新生が開始される際に、内皮細胞からAng1のアンタゴニストであるAng2が放出され、Tie2の一時的な不活性化により、壁細胞が離脱して血管新生が開始する。血管新生の開始において、まず糸状仮足を発現する先端細胞(tip cell)が発芽する。この細胞は遊走物質へ向かって移動して、血管の伸長方向をガイドし、その後方から増殖活性の高い、茎細胞(stalk cell)が血管伸長をになう。VEGFR-2の活性化した既存血管の内皮細胞の中で、Dll4(delta-like ligand 4; Notch受容体の結合因子)の発現、分泌の高まった細胞がtip細胞となり、tip細胞周囲の内皮細胞のNotchを活性化して、VEGFR-2やVEGFR-2とヘテロダイマーを形成してVEGFのシグナルを強める機能を有するneuropilin-1の発現を減弱させる(側方抑制)。このことで、tip細胞とは異なる表現型のStalk細胞が生じ、stalk細胞ではVEGF-Aに対する応答性が減弱し、Dll4の発現が減少する。そのために、tip細胞ではNotchの活性化が生じず、stalk細胞とはならずにtip細胞に分化決定する。
最終的には可溶性VEGF受容体1(sFlt1)の発現が高いファランクス細胞(phalanx cell)がVEGFを中和して血管新生を終了化し、壁細胞が裏打ちした安定血管の形成を誘導する。
図表
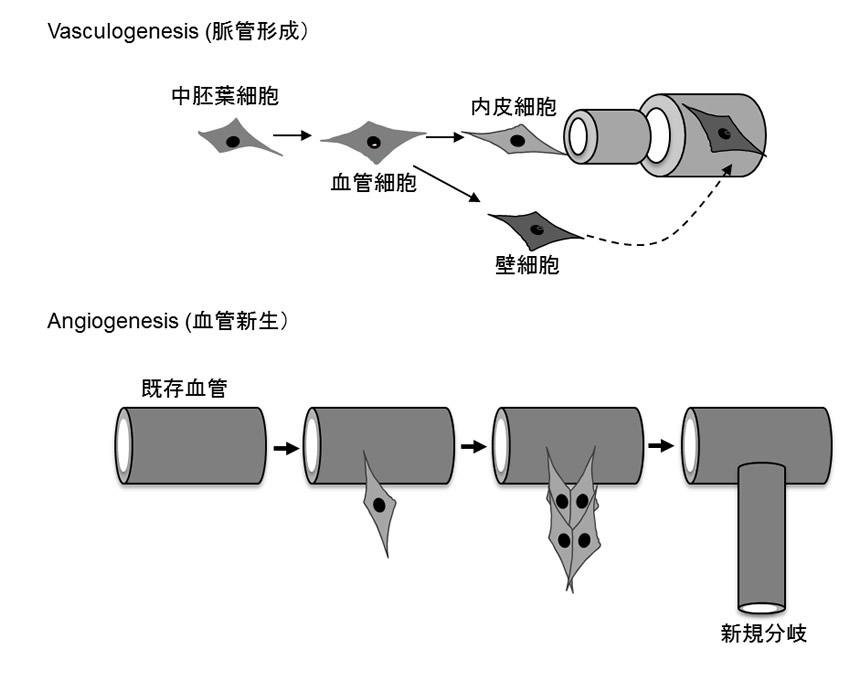
図1
参考文献
1) Phng LK, Gerhardt H: Angiogenesis: a team effort coordinated by notch. Dev Cell 16: 196